胆汁酸-肠道菌群在肠道炎症和肿瘤生成中的互作
综述 | 胆汁酸-肠道菌群在肠道炎症和肿瘤生成中的生理学机制

文献导读
胃肠道癌症的发病机制涉及到广泛且复杂的基因-环境相互作用。人类胃肠道中寄居着种类多样的共生细菌以及其它微生物,统称为微生物区系。宿主的基因组和微生物的基因组共同产生各种各样的代谢物,可以作为重要的信号分子和能量物质。这些动态集合的小分子代谢物,也称为代谢组, 与宿主的互作可调控免疫系统,代谢表型,增加罹患各种疾病的风险以及影响对治疗方法的响应。越来越多的证据发现肠道菌群可以促进肿瘤生成。一个常见的例子就是幽门螺杆菌诱导的胃癌;幽门螺杆菌通过产生毒力因子,细胞毒素相关基因A(CagA)的分泌引发胃萎缩和胃酸过少,这使得胃易受其他对pH敏感的细菌物种(如不动杆菌)过度生长的影响,从而导致细菌将膳食硝酸盐转化为致癌物的速度增加。在众多由菌群-宿主共代谢产生的一系列内源性代谢物中,胆汁酸因为可以促进肿瘤形成的特性而逐渐受到关注。这篇综述文章主要讨论胆汁酸的合成,代谢和转运,以及与肠道炎症和肿瘤生成的关系。
文章简介:
原标题:Bile acid–microbiota crosstalk in gastrointestinal inflammation and carcinogenesis
译名:胆汁酸-菌群在胃肠道炎症和肿瘤发生中的互作
期刊:《Nature Reviews Gastroenterology & Hepatology》
IF: 16.990
通讯作者:贾伟
擅长领域:「中药新制剂与新剂型、复方中药物质基础和配伍机制、中药代谢组学 」
通讯地址:上海市糖尿病重点实验室,上海交通大学附属第六人民医院
摘要
越来越多的证据表明,肠道微生物与罹患胃肠癌(如结直肠癌(CRC)和肝细胞癌(HCC))的风险,发展和进程之间存在着密切关系。在肝脏中产生的胆汁酸经由肠道微生物产生的酶来代谢产生种类丰富的胆汁酸库,胆汁酸库对维持健康的肠道微生物组成,平衡的脂质和碳水化合物代谢,胰岛素敏感性和先天免疫均发挥着重要作用。考虑到胆汁酸信号传导的复杂性以及肠道微生物与宿主之间的一个直接的生物化学相互作用,研究人员从系统生物学的角度来理解并阐明
肝脏-胆汁酸-微生物轴及其在胃肠癌发生中的作用以逆转由微生物介导的在疾病状态下发生的胆汁酸代谢类型的改变。所以需要对该领域最近的研究进展进行一个筛查与回顾。在本综述中,作者详细讨论了CRC和HCC中胆汁酸与胃肠癌发生之间的生物学机制,其涉及两种主要的胆汁酸受体,法尼醇X受体(FXR)和G蛋白偶联胆汁酸受体1(TGR5)。作者强调了在癌症治疗背景下靶向依赖于肠道-微生物的胆汁酸代谢改变的策略和先进技术。
主要内容
由于本篇文章是一篇内容十分详实的综述,大部分内容和以前分享过的关于胆汁酸合成,信号通路等内容有重复,关于这部分内容可以点击文末链接[1][2]进行再次阅读。另外由于本篇综述的图片内容非常精美,图例文字非常高度且周密的概括了文章中的文字部分内容,所以我将按照文章中的5幅图片按顺序进行翻译整理,供大家讨论学习。
一图看懂
1. 胆汁酸合成,转运和代谢

胆汁酸合成、转运和代谢。哺乳动物体内含量最高的两种初级胆汁酸为胆酸CA和鹅脱氧胆酸CDCA,对应的次级胆汁酸为脱氧胆酸DCA和石胆酸LCA。胆汁酸在肝脏中经由细胞色素P450介导的胆固醇的氧化而合成,包括经典和替代途径。啮齿动物肝脏中CDCA会转化为α-鼠胆酸α-MCA和β-鼠胆酸β-MCA。在猪中,CDCA主要通过CYP3A4的C-6α-羟化作用转化为猪胆酸HCA。然而在人中,主要为CA和CDCA。在肝脏中,胆汁酸主要通过bile acid-CoA synthetase(BACS)和bile acid-CoA:amino acid N-acyltransferase(BAAT)和甘氨酸和牛磺酸耦合,然后再通过胆盐输出泵(bile salt export pump, BSEP)运送到胆囊中储存。同时,肝脏也能通过磺基转移酶和UDP-葡糖醛酸转移酶作用生成磺酸化和葡糖醛酸化的胆汁酸,然后再与甘氨酸和牛磺酸耦合,通过多药耐药相关蛋白2(MRP2)转运并分泌到胆囊中。在小肠中,耦合形式的胆汁酸被胆酸盐水解酶BSH水解变成游离形式的胆汁酸,紧接着又通过7-α脱羟基作用生成次级胆汁酸。CA和CDCA分别转化为DCA和LCA,αT-MCA和Tβ-MCA经BSH生成游离的α-MCA和β-MCA。β-MCA通过C-6差向异构化生成ω-MCA,ω-MCA接着通过7α-脱羟基作用生成猪脱氧胆酸HDCA。CDCA还可以通过羟基类固醇脱氢酶HSDH形成熊去氧胆酸UDCA。葡糖醛酸,磺酸耦合的胆汁酸主要通过MRP2分泌到尿液中。参与BA代谢的主要肠道菌群包括具有解耦合作用的拟杆菌,梭菌,乳酸杆菌,双歧杆菌和李斯特菌; Bacteroides,Eubacterium,Clostridium,Escherichia,Eggerthella,Eubacterium,Peptostreptococcus和Ruminococcus能使C3,C7和C12的羟基氧化和差向异构化; 梭菌和真细菌具有7-脱羟基作用; 拟杆菌,真细菌和乳酸杆菌具有酯化作用; 梭菌,梭杆菌,球菌和假单胞菌具有脱硫作用。在回肠末端,大多数游离状态的BA(包括CA,CDCA,DCA,UDCA,HDCA,α-MCA,β-MCA和ω-MCA)可以被顶端钠依赖性BA转运蛋白(ASBT)重吸收进入肠细胞并通过基底外侧BA转运蛋白有机溶质转运子亚基α(OSTα),OSTβ,MRP2和MRP3分泌进入门静脉循环。 然后通过钠/牛磺胆酸盐共转运多肽(NTCP)和有机阴离子转运多肽1(OATP1)将BA吸收到肝细胞中。 肝脏MRP3,MRP4和OSTα-OSTβ复合物也为BA进入体循环提供了替代排泄途径。
2. 正常生理状态下和失调及炎症状态下的胆汁酸肝肠循环

正常生理状态下和肠道失调及炎症状态下的胆汁酸肝肠循环。a图表示的是正常生理条件下的肠道和肝脏之间的肝肠循环;b图展示的是肠道菌群失调和炎症状态下的肝肠循环过程。在肠道处于炎症状态下,肠道屏障功能会发生紊乱,从而降低肠道法尼醇X受体FXR的表达,导致成纤维细胞生长因子19(FGF19)-FGF受体4(FGFR4)信号通路活性降低,肠道胆汁酸结合蛋白(IBABP)下调和BA外排转运蛋白有机溶质转运蛋白亚基α(OSTα)和OSTβ的表达下调以及顶端钠依赖性BA转运蛋白(ASBT)上调。IBABP载体蛋白表达降低会导致从肠上皮细胞转移到OSTα-OSTβ外排复合体的胆汁酸减少,从而进入门静脉的胆汁酸也减少,然后破坏正常的肝肠循环。这一系列事件的发生,导致进入肠细胞的胆汁酸越来越多,但是却阻止其进入门静脉,所以积聚在肠上皮细胞的胆汁酸越来越多,最终导致肠道产生炎症。FGF19-FGFR4轴信号传导的减少会通过降低JUN-细胞外信号调节激酶(ERK)-细胞色素P450 7A1(CYP7A1)轴来增加肝脏中胆汁酸合成。肝脏炎症可以由肝脏细胞膜的胆汁酸紊乱引起,随后促炎症通路的激活会激活核因子kappa-B(NF-κB),NF-κB可以抑制肝脏FXR和进一步阻止小异二聚体伴侣分子SHP的激活。我们都知道,SHP的激活可以抑制胆汁酸合成过程中的限速酶CYP7A1的合成。所以这里FXR表达的降低会导致SHP的失活和CYP7A1水平的下降以及胆汁酸的流入会因为OSTα,OSTβ,MRP3和MRP4以及所有的小管转运蛋白而受到FXR的转录调控和促进胆汁酸的合成。依赖NF-κB的FXR转录抑制也能导致钠/牛磺胆酸盐共转运多肽(NTCP)的表达降低,其参与BA流入过程。然而,多特异性有机阴离子转运蛋白(OATP)不受FXR的影响。FXR还控制BA解毒作用,FXR的转录降低导致过氧化物酶体增殖物激活受体α(PPARα)及其靶基因的表达下降,其编码细胞色素P450酶(CYP),磺基转移酶(SULT)和UDP-葡糖醛酸基转移酶(UGT)。PPARα表达降低还会诱导多药耐药蛋白2(MDR2),MDR3,MRP3和MRP4转运蛋白表达的降低。在这些条件下,胆汁淤积和炎症都会加剧,这可能导致肝癌的发展。胆汁淤积通过各种机制引起炎症,导致NF-κB的上调和激活,然后NF-κB直接与FXR启动子结合以抑制其转录。
3. 胆汁酸诱导肝脏炎症和肿瘤生成
 胆汁酸诱导肝脏炎症和肿瘤生成。由于胆汁酸的亲脂性,洗涤剂性质,BA可以直接破坏质膜并引起蛋白激酶C(PKC)的活化,然后蛋白激酶C激活p38促分裂原活化蛋白激酶(p38MAPK)。由此激活p53和核因子kappa-B(NF-κB)而导致诱导细胞凋亡和炎症的增加。激活后的NF-κB易位至细胞核并促进编码促炎细胞因子如TNF,IL-1β和IL-6的基因转录,这些细胞因子可以正向激活NF-κB,从而形成炎症循环。IL-6可以激活Janus激酶(JAK)-信号转导和转录激活因子3(STAT3)途径,导致细胞凋亡降低和肝细胞癌(HCC)的进展。IL-1β还激活磷酸肌醇3-激酶(PI3K)-MDM2通路以反向调节p53,从而通过减少细胞凋亡来增加DNA受损细胞的存活,这可能导致HCC。还显示NF-κBp65亚基直接与FXR结合并抑制其转录活性,导致胆汁酸转运蛋白(如有机溶质转运蛋白α(OSTα),OSTβ,胆盐输出泵(BSEP),多药耐药相关蛋白2(MRP2),多药耐药蛋白(MDR2)和MDR3)的表达降低,导致胆汁酸外流减少和胆汁淤积,以及胆汁酸生物合成增加。FXR转录的抑制导致肝脏中胆汁酸水平增加从而导致HCC的炎症。 胆汁酸通过干扰细胞膜也可以激活磷脂酶A2(PLA2),导致细胞膜释放花生四烯酸(AA),AA可被肝脏中的环氧合酶(COX)和脂氧合酶(LOX)代谢产生活性氧(ROS),并因此诱导DNA损伤。ROS可直接激活NF-κB,也可直接诱导细胞DNA损伤,从而导致HCC
胆汁酸诱导肝脏炎症和肿瘤生成。由于胆汁酸的亲脂性,洗涤剂性质,BA可以直接破坏质膜并引起蛋白激酶C(PKC)的活化,然后蛋白激酶C激活p38促分裂原活化蛋白激酶(p38MAPK)。由此激活p53和核因子kappa-B(NF-κB)而导致诱导细胞凋亡和炎症的增加。激活后的NF-κB易位至细胞核并促进编码促炎细胞因子如TNF,IL-1β和IL-6的基因转录,这些细胞因子可以正向激活NF-κB,从而形成炎症循环。IL-6可以激活Janus激酶(JAK)-信号转导和转录激活因子3(STAT3)途径,导致细胞凋亡降低和肝细胞癌(HCC)的进展。IL-1β还激活磷酸肌醇3-激酶(PI3K)-MDM2通路以反向调节p53,从而通过减少细胞凋亡来增加DNA受损细胞的存活,这可能导致HCC。还显示NF-κBp65亚基直接与FXR结合并抑制其转录活性,导致胆汁酸转运蛋白(如有机溶质转运蛋白α(OSTα),OSTβ,胆盐输出泵(BSEP),多药耐药相关蛋白2(MRP2),多药耐药蛋白(MDR2)和MDR3)的表达降低,导致胆汁酸外流减少和胆汁淤积,以及胆汁酸生物合成增加。FXR转录的抑制导致肝脏中胆汁酸水平增加从而导致HCC的炎症。 胆汁酸通过干扰细胞膜也可以激活磷脂酶A2(PLA2),导致细胞膜释放花生四烯酸(AA),AA可被肝脏中的环氧合酶(COX)和脂氧合酶(LOX)代谢产生活性氧(ROS),并因此诱导DNA损伤。ROS可直接激活NF-κB,也可直接诱导细胞DNA损伤,从而导致HCC
4. 胆汁酸诱导巨噬细胞的TGR5信号通路

胆汁酸诱导巨噬细胞的TGR5信号通路。巨噬细胞分为M1和M2两种类型,M1巨噬细胞是促炎性的,M2细胞则具有免疫抑制。尽管胆汁酸(BAs)不能诱导全部的巨噬细胞极化成M1或M2细胞表型,但它们已被证明可激活G蛋白偶联胆汁酸受体1(TGR5)并产生出一种主要表现为免疫抑制的“混合细胞表型”,因为IL-10:IL-12比值增加了。TGR5可以激活腺苷酸环化酶-cAMP和表皮生长因子受体(EGFR)-SRC激酶(SRC)途径。在促炎性M1样通路(虚线箭头)中,EGFR的TGR5依赖性反式激活可通过激活SRC诱导促炎信号,SRC反过来激活细胞外信号调节激酶1/2(ERK1/2)和蛋白质激酶C(PKC)。当EGFR和相关的G蛋白偶联受体(GPCR)如TGR5在脂筏中并排共存(由胆固醇和鞘脂标识)时,已经显示出这种反式激活。 在TGR5激活后,发生EGFR配体肝素结合EGR(HB-EGF)的金属蛋白酶依赖性切割,从而激活EGFR-SRC途径。PKC的TGR5依赖性激活导致核因子κ-B(NF-κB)的激活,其导致促炎细胞因子IL-1β,IL-6和TNF的表达增加,以及ERK1/2的进一步自分泌激活和持续的促炎信号循环。M1样表型中ERC1/2的SRC依赖性激活导致MYC的表达增加,其增加细胞凋亡。促炎M1样信号通过其产生的促炎细胞因子调节先天免疫,特别是通过增加促炎反应型17T辅助细胞(TH17)的活化(响应增加的IL-1β,IL-6)水平和TH1细胞(响应增加的IL-12,IFNγ水平)同时抑制免疫抑制性T调节(Treg)细胞(响应IL-10水平降低)。在BA依赖性激活TGR5期间,促炎性M1样表型占主要。同时,SRC激活的同时也激活信号转导和转录激活因子3(STAT3),其促进抗炎M2样作用(实线箭头),包括TH17和TH1细胞活化减少,TNF,IFNβ,IL-6和 IL-12和IL-10和转化生长因子β(TGFβ)的产生增加,这与IL-10:IL-12比率的增加相对应。BA激活的TGR5也独立于EGFR反式激活而激活cAMP途径; cAMP激活蛋白激酶A(PKA),其导致cAMP反应元件结合蛋白(CREB)表达和活性的上调。FOS是CREB发挥活性的重要靶基因产物,其与活化的NF-κB的p65亚基结合以抑制其向细胞核的易位,是TGR5发挥抗炎作用的一个方面。CREB活性的另一个重要靶基因产物是IL-10,其发挥免疫抑制作用因此具有抗炎作用
5. 肠道中的胆汁酸影响结直肠癌
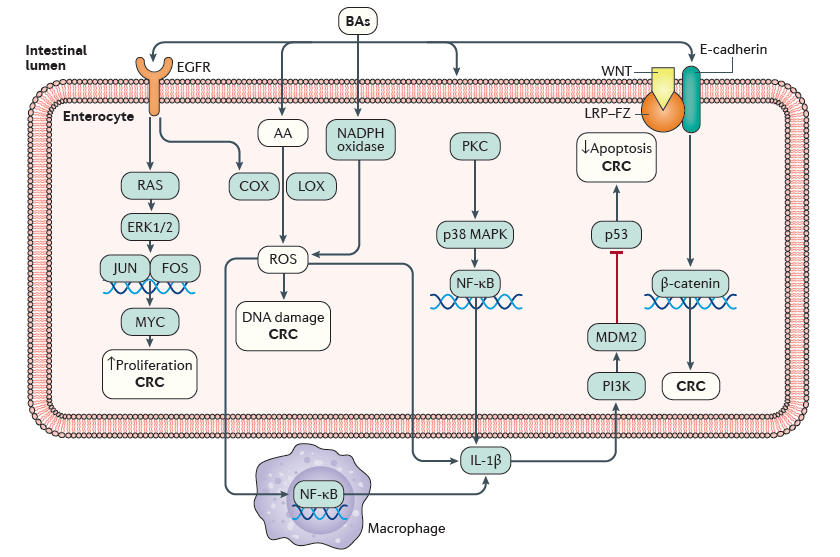
肠道中的胆汁酸影响结直肠癌CRC。次级胆汁酸,尤其是DCA,可以影响肠细胞的一些信号通路从而导致结直肠癌的发生。BA可以激活表皮生长因子EGFR,EGFR可以导致RAS-细胞外信号调节激酶1/2(ERK1/2)信号传导的激活。ERK1/2诱导JUN和FOS的激活和核转位,它们共同形成激活蛋白1(AP-1)转录因子复合物,促进AP-1靶基因MYC210的转录。 MYC的表达可以增加细胞增殖和多能性。 EGFR的激活还导致环加氧酶(COX)和脂氧合酶(LOX)的上调,其促进活性氧(ROS)的产生。BA可以破坏质膜以释放花生四烯酸(AA)进入细胞质,在细胞质中AA可以迅速用作COX和LOX生成ROS的底物。NADPH氧化酶可被胆汁酸刺激,从而进一步增加ROS的产生。在肠上皮细胞中,ROS可引起DNA损伤和炎症,导致促炎细胞因子IL-1β的表达上调。以上这些变化都会有助于CRC的发生和进展。胆汁酸对质膜的扰动也激活蛋白激酶C(PKC),随后PKC激活p38促分裂原活化蛋白激酶(p38MAPK)和核因子-κB(NF-κB)。 处于激活状态的NF-κB易位到细胞核中,并促进IL-1β的转录。 IL-1β可以以自分泌和旁分泌方式发出信号来激活磷脂酰肌醇3-激酶(PI3K)-MDM2轴,从而导致p53活性的阻断。 p53活性降低导致细胞凋亡减少和DNA受到损伤的细胞存活率增加,如此一来更加有利于CRC的发展的可能性。另外,胆汁酸还可以通过未知的机制来刺激WNT与低密度脂蛋白受体相关蛋白(LRP)-卷曲受体(FZ)-E-钙粘蛋白-β-连环蛋白复合物的结合,从而导致β-catenin从E-钙黏蛋白中释放后进入细胞质,再从细胞质被转移至细胞核并刺激转录因子,如T细胞因子(TCF)-淋巴增强因子(LEF),以增加细胞增殖和细胞干性,促进CRC的发生
靶向胆汁酸信号通路
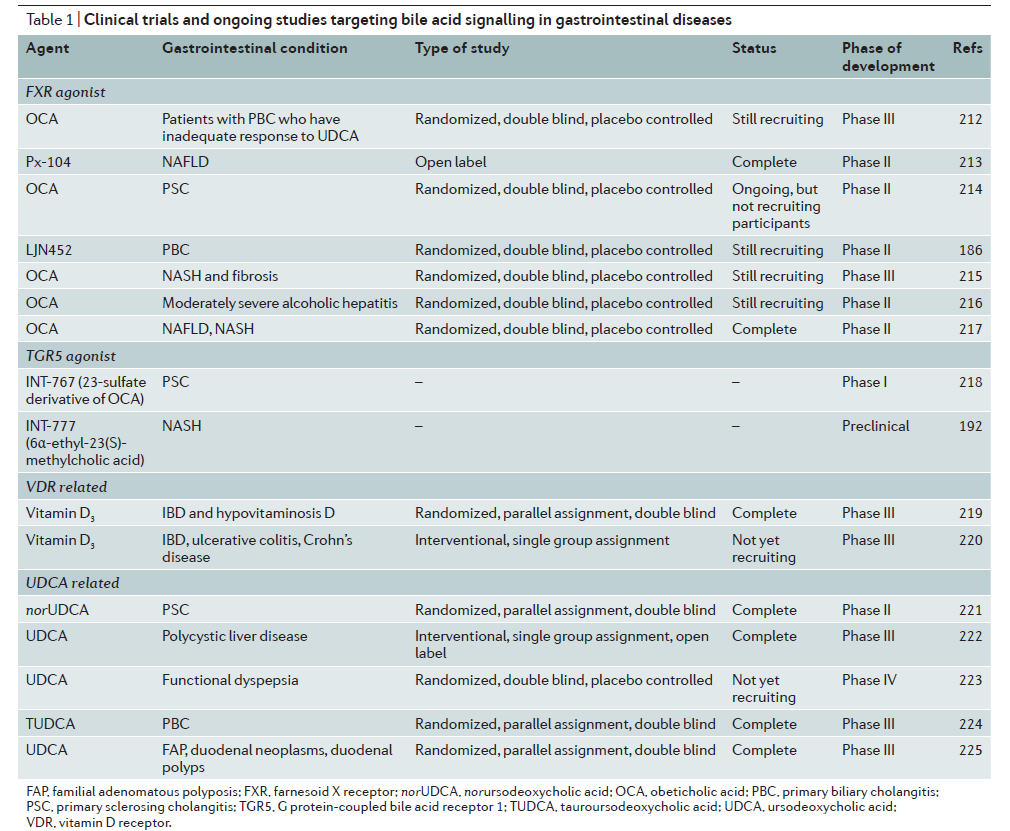
根据前面的内容我们知道,胆汁酸信号通路诸如FXR和TGR5的紊乱会导致胆汁酸肝肠循环的紊乱,从而导致肠细胞胆汁酸持续增多,并进一步导致肠道炎症,发生CRC或HCC。所以靶向FXR和TGR5信号通路对于恢复肝肠循环正常化和控制肠道炎症可能是一种有效的方法。表1(TABLE 1)总结了靶向肠道疾病的胆汁酸信号的研究进展。
总结
除了促进膳食脂肪的吸收外,胆汁酸通过不同的核受体和细胞信号传导途径来作为一种强信号分子,以调节脂质,葡萄糖和能量代谢。 过去三十年的研究对我们理解胆汁酸代谢机制和胃肠道癌症发病机制作出了重大贡献。 在本综述中,作者详细讨论了胆汁酸合成,转运和代谢的机制,并且强调了在正常和疾病状态下胆汁酸合成和转运的信号调节。回顾了胆汁酸,肠道微生物群,胃肠道炎症和癌症之间的联系,特别关注与CRC和HCC的联系。 最后,胆汁酸,胆汁酸激活的核受体(如FXR,PXR,CAR和VDR)和TGR5是未来开发治疗胃肠癌的药物的关键治疗靶点。
展望
如果我们能够很好的理解胆汁酸对其受体的激素效应,那么就可以比较好的预防和控制经常表现出性别差异的胃肠癌症的发生。并且通过理清胆汁酸合成,转运和代谢中涉及的通路调节的生理学基础,从而通过控制胆汁酸的数量和类型来预防肠道和肝脏中的胆汁酸毒性。
参考
[1] https://mp.weixin.qq.com/s/yk7MtaeQWasvFp-pZD7AWw
[2] https://mp.weixin.qq.com/s/luo7iu3RDJ0jQXtoQ3XoTg
[3] 原文链接
深度基因小伙伴温馨提示:
- 如果我们对文章理解有偏差,非常欢迎大家向我们反馈,我们会认真阅读建议并修改,另外有意愿加入我们的小团队的老师和同学可发送邮件至我们的邮箱:deepgener@gmail.com 祝大家科研顺利,生活开心!
- 想要了解更多内容请访问我们的深度基因网站:http://deepgener.wordpress.com/
- 点击查看上一篇文章
胆汁酸-肠道菌群在肠道炎症和肿瘤生成中的互作相关推荐
- Nature:微生物的胆汁酸代谢物调节宿主肠道特异Treg细胞的稳态
撰文 | 崔浩白 责编 | 兮 人体肠道微生物可代谢多种宿主或食物来源的前体分子,从而产生大量具有生物活性的下游代谢产物.宿主与肠道微生物之间的胆汁酸代谢网络(biliary network)便是其中 ...
- 什么是胆汁酸,其与肠道微生物互作如何影响人体健康
谷禾健康 胆汁酸是一种代谢胆固醇的物质,它具有两个亲和性不同的区域,即一部分能够与水分子相互作用(亲水),而另一部分则不能与水分子相互作用(疏水). 由于拥有这种两亲性质,胆汁酸能够在生物体内与胆固醇 ...
- 肠道菌群检测在临床感染判别中的应用
谷禾健康 感染是人类面临的健康威胁之一.各种病原体,如细菌.病毒.真菌.寄生虫等,存在于我们日常接触的环境.物品.食物等中.一些常见的感染病例包括感冒.流感.腹泻.组织器官或血液感染等,在全球范围内广 ...
- 常见水果对肠道菌群、肠道蠕动和便秘的影响
便秘的全球患病率为14%.慢性便秘会影响生活质量.健康人相比,便秘还会导致粪便重量降低,且与结直肠癌风险增加相关. 饮食调整是治疗便秘的主要方法之一.世界胃肠病学协会建议通过饮食建议或补充来增加纤维摄 ...
- 如何获取瘦人肠道菌群_肠道菌群移植—让你获得窈窕身材
原标题:肠道菌群移植-让你获得窈窕身材 "遍寻不着,犹叹当年小蛮腰,空余恨,如今一身五花膘.". 不管是曾经瘦过,还是现在胖着,人们对窈窕身材的向往和追求从未停止过. 肥胖是什么? ...
- iMeta | 中科院李小方等膳食甘草促进小鼠镉解毒并调节肠道菌群代谢
点击蓝字 关注我们 iMeta | 膳食甘草促进小鼠镉解毒并调节肠道菌群代谢 https://doi.org/10.1002/imt2.7 iMeta Volume 1 Issue 1 2022.3 ...
- 猫狗肠道菌群—“主子们”的健康新领域
随着生活水平提升,我国养宠人士逐年增多.数据显示,我国城镇养宠人士达到6844万多人,且仍在不断增长. 其中养猫人群数占比59.5%,养狗人群数占比51.7%,养水族类占比8.3%,养爬行类宠物的占比 ...
- iMeta | 调控肠道菌群的宿主源代谢分子概述
点击蓝字 关注我们 调控肠道菌群的宿主源代谢分子概述 iMeta主页:http://www.imeta.science 研究论文 ● 原文链接DOI: https://doi.org/10.1002/ ...
- 阳康后是否二次感染,长新冠与肠道菌群的关联,多种潜在的干预措施
谷禾健康 随着大家陆续"阳康",大家逐渐恢复以往的生活,城市的烟火气回来了. 然而阳康后真的万事大吉了吗? 还是有很多朋友处于这样的状态:感觉恢复了,又好像没有完全恢复,身体多少有 ...
最新文章
- DOS批处理高级教程精选(六)
- 明晚8点直播 | Transformer新型神经网络在机器翻译中的应用
- C#开发Unity游戏教程之Unity中方法的参数
- python画圣诞树代码-圣诞节!教你用Python画棵圣诞树
- session无法传值解决方案!
- [译]JavaScript:ES6中的模板字符串简介
- 经典算法题每日演练——第七题 KMP算法
- Talos实验室深入我国DDoS黑市DuTe 揭露各种DDoS团伙、平台、工具及攻击
- 读写生信流程必备的 Perl 语法
- 第4章 最基础的分类算法-k近邻算法 kNN 学习笔记 下
- 什么是低代码(LowCode)/无代码(NoCode)平台?
- 曾宪武《物联网通信技术》课后答案(三)
- 疯狂模渲大师体验版安装教程|效果图设计师怎么安装并注册3dmax疯狂模渲大师体验版?
- Windows 10出现0x8000ffff错误应该如何解决?
- DocLocker - 文档外发控制系统
- xp计算机无法正常启动,修复XP系统无法正常启动进入不了计算机的解决办法
- java乐视面试题_java面试题经典解读
- shipyard安装不迷茫
- 准确的找到BAT实习机会~我入职了腾讯
- Python 实现个人博客系统